I am an NSF postdoctoral scholar in the van Breukelen laboratory at the University of Nevada Las Vegas. I am a comparative physiologist fundamentally interested in the evolution of endothermy. There are numerous robust hypotheses or models to explain the evolution of endothermy/homeothermy in mammals. 1) The Body-Size Reduction Hypothesis predicts that as therapsids (mammalian ancestors) decreased in size, there was a required increase in metabolism. Earlier therapsids were generally large, typically 20-100 kg. As a result, these animals had greater thermal inertia and more constant body temperature in which the molecular machinery for metabolism operated. The model predicts reduction in size required increases to metabolism to account for surface area to volume ratios and still maintain warmer and more constant temperature. 2) The Niche Expansion Hypothesis predicts increased metabolism allowed mammals to occupy new niches e.g., nocturnal activity patterns. 3) The Aerobic Capacity Model argued that elevated metabolic rates facilitated sustained locomotor activities. 4) The Parental Care Model suggested warmer body temperatures allowed for greater resource allocation to offspring. 5) The Assimilation Capacity Model was similar in that warmer body temperatures enhanced digestive efficiencies and allowed for greater resource transfer to offspring.
It is important to note that these and any other hypothesis for the evolution of endothermy may not be mutually exclusive. It is also important to note that both the body-size reduction hypothesis and niche expansion hypothesis are difficult to directly test. Most attempts to address the veracity of the models have utilized models such as mice or rats where body temperature and metabolism are linked and very constrained e.g., fluctuation of core body temperature is minimal. The previous challenge for directly testing hypotheses related to the evolution of endothermy has been the unavailability of an appropriate mammalian model system wherein fluctuating body temperature is not accompanied by a marked metabolic depression. A model that exhibits fluctuating body temperatures without the expected associated change in metabolism would be perfect for the direct testing of models related to the evolution of endothermy. Tenrecs (Tenrec ecaudatus) are a perfect model to untangle the selective pressures that may have given a rise to the increased endothermy in “modern mammals”.
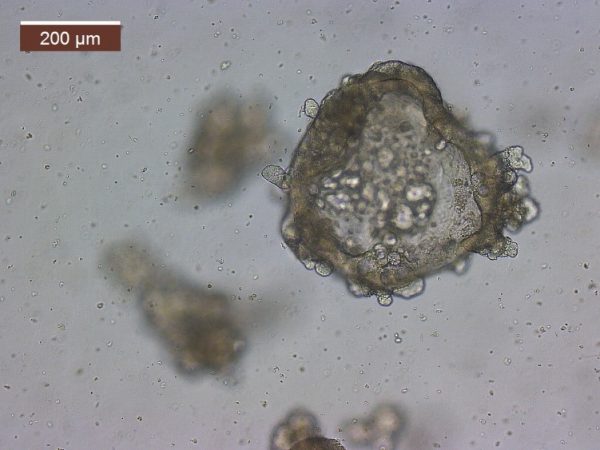
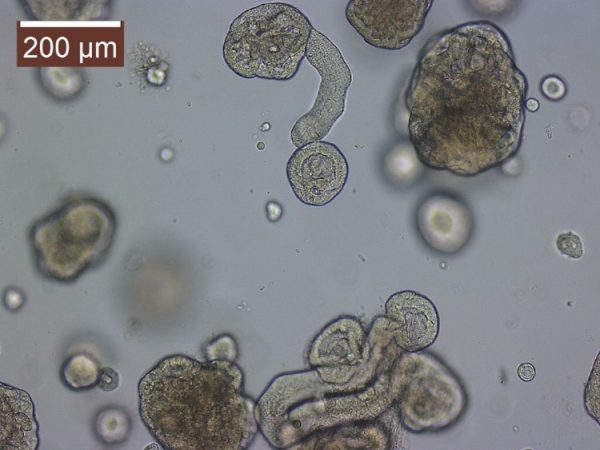
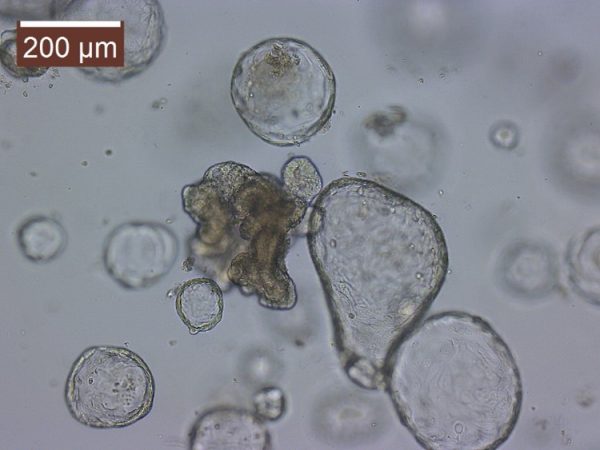
My current NSF grant specifically funds the investigation of the Assimilation Capacity model. So fundamentally I am investigating the influence of body temperature on digestive efficiency. The van Breukelen laboratory become acquainted with the 3D cell culture technique referred to as organoids. Organoids are a multicellular culture system that more recapitulates the native tissue in which it was derived from. So essentially intestinal organoids can possess functional enterocytes, goblet cells, and paneth cells. We become aware of the Drs. Mochel and Allenspach-Jorn lab at Iowa state university, developing organoids in non-model organisms. So, we believed they would be perfect collaborators.
I worked closely with PhD student Chris Zdyrski and with these collaborations, I have developed an in vitro 3D cell culture system from tenrec jejunum and tenrec /golden mantled ground squirrel kidney tissue. Without the funding from the RCN, I would never have gotten the opportunity to learn such a novel/innovative technique. We are currently in the process of characterizing these organoids to assess the presences of functional transporters. I hope that the system will allow me a future opportunity (as an independent investigator) to address mechanistic questions that are more difficult to ask with a simple cell isolation e.g. I can address the role of pH regulation, hypoxia, temperature, etc. on tissue metabolism. Again, I am thankful for the RCN lab exchange and to everyone at Iowa State University.